什么是杂种优势请介绍杂种优势的遗传学理论,你认为这些理论完满吗你的看法如何有人建议用无融合生殖来固定植物的杂种优势,你觉得可行吗为什么(提示:无融合生殖是指植物不通过受精产生胚和种子的生殖方法。)
参考答案:
解析:杂种优势(heterosis)是生物界的普遍现象。它是指两个遗传组成不同的亲本杂交产生的杂种第一代,在生长势、生活力、繁殖力、抗逆性、产量和品质上比其双亲优越的现象。杂种优势所涉及的性状大都为数量性状,故必须以具体的数值来衡量和表明其优势的程度。但也有人认为F1的性状表现必须超过其双亲中最优的亲本,才算是真正具有杂种优势。
杂种优势的表现是多方面的,而且是很复杂的。按其性状表现的性质,大致可以分为三种类型:一是杂种营养体发育较旺的营养型;二是杂种生殖器官发育较盛的生殖型;三是杂种对外界不良环境适应能力较强的适应型。这三种类型的划分只是相对的,实际上它们的表现总是综合的,这里不过是按其优势明显的性状作为一个划分的标志。
关于杂种优势的遗传解释主要有显性假说和超显性假说:
1)显性假说:1910年布鲁斯(A. B. Bruce)等人首先提出显性基因互补假说,1917年琼斯(D. F. Jones)又进一步补充为显性连锁基因假说。这一假说简称为显性假说(dominance hypothesis)。
显性假说认为杂种优势是由于双亲的显性基因全部聚集在杂种中所引起的互补作用。基柏(F. Keeble)和皮洛(C. Pellow)于1910年最早提出这方面的试验论证,他们曾以两个株高5~6英尺的豌豆品种进行杂交试验,一个品种的茎秆是节多而节间短,另一个品种的茎秆是节少而节间长,其F1聚集了双亲的节多和节间长的显性基因,因而株高达到7~8英尺,表现出明显的杂种优势。
根据显性假说,按独立分配规律,如所涉及的显隐性基因只是少数几对时,其F2的理论次数应为(3/4+1/4)n的展开,表现为偏态分布。但是,事实上F2一般仍表现为正态分布。F2以后虽然优势显著降低,但在理论上应该能从其后代选出像F1同样优势,并且把全部纯合显性基因聚合起来的个体。而事实上很难选出这种完全纯合显性基因的后代。为此,显性连锁基因假说作了补充的解释,认为一些显性基因与另一些隐性基因位于各个同源染色体上,形成一定的连锁关系。而且控制某些有利性状的显性基因是非常多的,即n很大时,则F2将不是偏态分布,而是正态分布了。同时,在这样非常大的分离群体中,选出完全纯合显性基因的个体也几乎是不可能的。
为了说明显性假说,现以玉米的两个自交系为例。假定它们有5对基因互为显隐性的关系,分别位于两对染色体上。同时假定各隐性纯合基因(如aa)对性状发育的作用为1,而各显性纯合和杂合基因(如AA和Aa)的作用为2。这两个自交系杂交产生的杂种优势可表示如下:
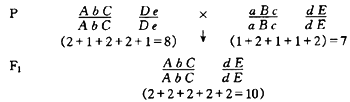
由此可见,由于显性基因的作用,F1比双亲表现了显著的优势。显性假说得到许多试验结果的验证。但是,这一假说也存在着缺点,它考虑到等位基因的显性作用,但没有指出非等位基因的相互作用,即上位效应。
2)超显性假说:超显性假说(overdominance hypothesis或superdominance hypothesis),也称等位基因异质结合假说。这个假说的概念最初是由肖尔(Shull)和伊斯特(East)于1908年分别提出的,他们一致认为杂合性可引起某些生理刺激,因而产生杂种优势。伊斯特于1936年对超显性假说作了进一步说明,指出杂种优势来源于双亲基因型的异质结合所引起的基因间的互作。根据这一假说,等位基因间没有显隐性的关系。杂合等位基因间的相互作用显然大于纯合等位基因间的作用。假定a1a1是一对纯合等位基因,能支配一种代谢功能,生长量为10个单位;a2a2是另一对纯合基因,能支配另一种代谢功能,生长量为4个单位。杂种为杂合等位基因a1a2时,将能同时支配a1和a2所支配的两种代谢功能,于是可使生长量超过最优亲本而达到10个单位以上。这说明异质等位基因优于同质等位基因的作用,即a1a2>a1a1,a1a2>a2a2。由于这一假说可以解释杂种远大于最优亲本的现象,所以称为超显性假说。
两个亲本只有一对等位基因的差异,杂交能出现明显的杂种优势,这是对超显性假说最直接的论证。某些植物的花色遗传是一对基因的差别,但它们的杂种植株的花色往往比其任一纯合亲本的花色都要深。例如,粉红色×白色获得F1表现为红色;淡红色×蓝色获得F1表现为紫色。而它们的F2不分离为简单的1:2:1的比例。
根据生化遗传学的许多试验资料,对于一对异质等位基因表现优势已得到一些有力的解释。例如,已知某些植物同一位点上两个等位基因能各自决定锈病的一个生理小种的抗性,因此,杂合体能抵抗两个生理小种,而纯合体只能抵抗一个生理小种。斯瓦尔茨(D. Schwartz)和洛夫纳(W. J. Laughner)于1969年曾研究酶活性的杂种优势,他们把一个不稳定而活泼的酶与一个稳定而不活泼的酶的异质等位基因结合起来,获得的杂种酶在活性上是既稳定而又活泼的。这些资料都表明一对异质等位基因的互作,常可导致来源于双亲的新陈代谢功能的互补或生化反应能力的加强等。
为了说明超显性假说,现同样假定玉米的两个自交系各有5对基因与生长势有关,各等位基因均无显隐性的关系。同时,假定a1a1、b1b1等为同质等位基因时的生长量为1个单位,而a1a2、b1b2等为异质基因时的生长量为2个单位。这两个自交系杂交产生的杂种优势可表示如下:
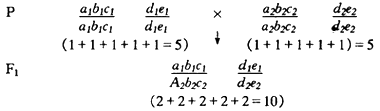
由此可见,由于异质基因的互作,F1的杂种优势可以显著地超过双亲。如果非等位基因间也存在互作,则杂种优势更能大幅度地提高。
由于非等位基因的互作而影响到性状的表现,称为上位性。费雪(R. A. Fisher)和马瑟(K. Mather)曾认为上位性在近亲繁殖和杂种优势的表现中,即使不是一个主要因素,也是一个重要因素。只是因为大多数性状都是受多基因控制的,在性状表现上等位基因的互作和非等位基因的互作一般是很难区分的。
普通小麦(Triticum aestivum)的起源常可作为说明非等位基因互作的一个突出的例证。普通小麦是由一粒小麦(Tr. monococcum)、拟斯卑尔脱山羊草(Ae. speltoides)和方穗山羊草[Ae. squarrosa,或称为节节麦(Tr. tauschii)]先后天然杂交形成的。这三个物种虽然都是没有生产价值的野生种,但是它们组成的普通小麦却在生产上具有极大的遗传潜力。因此,根据它们的遗传组成的比较分析,可以推论普通小麦在形成过程中所表现的杂种优势应该不是由于这三个野生种染色体内基因的互作,而是由于它们的染色体间基因的互作,也就是说,是由于非等位基因间互作的结果。当然,形成异源多倍体的几个原始物种,如果是来源于共同的祖先,那么它们的染色体之间的基因可能具有共同的位点,因而它们的杂种优势也可以说是由于等位基因互作的结果。所以,这些都进一步说明等位基因的互作与非等位基因的互作是不容易区分的。
越来越多的试验资料支持超显性假说。但是这一假说也存在着缺点,它完全排斥了等位基因间显隐性的差别,排斥了显性基因在杂种优势表现中的作用。并且,许多事实证明,杂种优势并不总是与等位基因的异质结合一致。例如,在自花授粉植物中,有一些杂种并不一定比其纯合亲本表现较优。
以上两个假说的共同点在于:都立论于杂种优势来源于双亲基因型间的相互关系。它们的不同点在于:前者认为杂种优势是由于双亲的显性基因间的互补;后者认为杂种优势是由于双亲等位基因间的互作。综观生物界杂种优势的种种表现,这两种假说所解释的情况都是存在的。所以,概括地说,杂种优势可能是由于上述的某一个或几个遗传因素造成的,即可能是由于双亲显性基因互补、异质等位基因互作和非等位基因互作的单一作用,也可能是由于这些因素的综合作用和累加作用。
必须指出,不论显性或超显性假说都只考虑到双亲细胞核之间的异质作用,完全没有涉及母本细胞质和父本细胞核之间的关系。事实上双亲杂交结合后,其F1的核质之间也可能存在一定的相互作用,因而引起杂种优势的表现。60年代以来,许多学者报道玉米、高粱、小麦、棉花等作物在亲本间的线粒体活性上所出现的互补作用,与其杂交产生的杂种优势表现具有密切的关联。因而有人提出利用两个杂交亲本间的线粒体活性互补作用,作为预测杂种优势的方法。这些试验结果都表明细胞质内的遗传物质及其与细胞核的相互作用,对于杂种优势的产生也是一个不可忽视的因素。
关于杂种优势的遗传机理,迄今都没有比较完善的解释。因此,这方面的遗传机理还需要通过生产实践和科学实验进一步的分析和研究。
根据上述杂种优势的遗传理论,已知杂种优势的来源在于F1的基因型的异质结合。因此,只有最大限度地保持F1基因型的杂合性,才能固定或部分固定杂种优势。因此,目前在固定杂种优势的研究上,一般认为比较有效的途径就是利用无性繁殖方法,使具有杂种优势的F1分生繁殖。通常甘薯和马铃薯等无性繁殖作物的杂交育种,就是利用F1的块根或块茎等繁殖固定其杂种优势的。现代生物科学中组织培养技术发展迅速,如果能够利用F1的体细胞,经过简便培养即能形成基因型一致的大量幼苗,那将可能完全省略年年大田制种。
利用无融合生殖的方法也将是一个比较有效的途径,由胚囊或珠心细胞发生无孢子生殖,形成二倍体胚和种子;或者由珠心组织在胚囊里形成二倍体的不定胚。这些胚和种子也将可以延续F1的杂合性,从而固定其杂种优势。