如何用遗传学方法(不是分子生物学方法)将基因定位到染色体上,请从人类和果蝇中各举一例予以说明。
参考答案:
解析:对人类来说,把家系分析和细胞学观察结合起来,可以发现某一性状的遗传与某一畸变染色体的传递有平行关系,由此把决定这一性状的基因定位在某一染色体的某一区域,作成细胞学图(cytological map)。例如有一家系中,红细胞型酸性磷酸酯酶1(acid phosphatase 1)活性的缺乏与2号染色体短臂的微小相关联,从而把酸性磷酸酯酶1基因(ACP 1)定位在2号染色体短臂的远端。
这几年来发展了一种新技术,可以绕过减数分裂过程,应用细胞培养方法,研究体细胞融合、突变、分离以及连锁和交换等,也就是用体细胞遗传学(somatic cell genetics,或简称cell genetics)方法,把基因定位在染色体上,作成细胞学图。
有一种病毒,如仙台病毒(Sendai virus)对细胞融合很有用处。病毒通常有一个特定附着点(attachment point),附着到宿主细胞上,并由此进入细胞。仙台病毒有几个附着点,如果两个细胞靠近在一起,它就能同时附着到两个不同的细胞。一个病毒比起细胞来要小得多,所以它所附着的两个细胞靠得很近,因而在某些情况下,两个细胞的膜可以融合。近年来也有用化学药剂聚乙二醇(polyethylene glycol,PEG)取代仙台病毒的。这种药剂可能使细胞膜部分降解,并在细胞间形成细胞质桥,从而提高细胞融合的效率。
如果把人体细胞和营养缺陷型的小鼠细胞(或仓鼠细胞等)混合培养,再加上促融因子——紫外线灭活的仙台病毒或PEG,那么两种细胞就有可能融合。融合细胞中有两个核,是异核体(heterokaryon)。异核体的两个核融合,形成杂种细胞。如果所用的小鼠细胞是营养缺陷型,那么要使这种细胞能够生长,非得在培养基上添加某种营养物不可。可是我们可通过细胞融合技术,把小鼠细胞和人体细胞融合,形成杂种细胞。杂种细胞含有小鼠染色体和人体染色体,小鼠的营养缺陷可由人体染色体上的有关基因的作用来弥补,所以培养基上不添加某种营养物质,杂种细胞也可保持下去。这种细胞往往有整套的小鼠染色体和丢失后保留下来的少许人体染色体,其中当然含有能补偿小鼠营养缺陷的那个染色体。通过不同的选择技术,再加上机遇性的变化,可以形成各种杂种细胞系,含有不同数目和不同号码的人体染色体。
上述这些过程,可以在显微镜下追循。这是因为近年来,染色体的染色技术有很大的发展,特别是应用了荧光染色法和其他特殊染色技术,已可使染色体纵长上呈现各种不同的分带(band)。这些分带的位置、宽窄和浓淡等随染色体号码的不同而不同。但就某一种分带技术来说,每一染色体的分带模式(banding pattern)是高度专一和恒定的。所以在杂种细胞中,非但小鼠染色体很容易跟人的染色体区分开来,而且人染色体的丢失过程也可追循,杂种细胞中留下来的人的染色体是哪条也比较容易认出。
这样我们就可以把人的基因定位在某一染色体上了。假定某人体细胞有一个或几个标记基因(gene marker),这些基因可以是控制营养需要或抗药性,也可以是控制细胞表面抗原或异常蛋白的形成等。我们实验的目的是要把其中一个或几个基因定位到特定的染色体上。
我们有不同的杂种细胞系,每个细胞系中除了小鼠染色体外,还有少数人的染色体。我们检验这些细胞系,把某一标记基因的在或不在与每一细胞中人的某一染色体的在或不在联系起来,从而推断某一基因是在某号染色体上。
在不同的杂种细胞系中,例如基因1和3或一起出现,或共同不见,所以我们可以下结论说,这两基因是连锁的。还有,基因1和3的在或不在直接跟第二染色体的在或不在有关,所以我们可以认为这两基因是同线的,都在第二染色体上。
用这种方法,已有相当数目的基因被定位于特定的染色体上。可是还不能像连锁群那样,把基因的顺序和基因间的距离都推算出来。这还有待于其他技术,例如某一染色体缺少了小小的一段,某个标记基因就不存在了,这样我们就知道这个基因就在缺失了的这一段上。例如上面提过的酸性磷酸酯酶基因ACP1就是通过体细胞杂交技术和染色体微小缺失方法而被进一步正确地定位于2号染色体短臂2带3区(2p23)的。
利用伴性遗传可以将果蝇的基因定位到染色体上。例如果蝇的野生型眼色都是红色,但是摩尔根在研究的早期发现一只雄蝇,复眼的颜色完全白色,这只白眼雄蝇与通常的红眼雌蝇交配时,子一代不论雌雄都是红眼,但子二代中雌的全是红眼,雄的半数是红眼,半数是白眼。如果雌雄不论,则子二代中3红眼:1白眼。这显然是个孟德尔比数,但与一般孟德尔比数不同之点是,白眼全是雄蝇。
另外,摩尔根也做了回交试验。最初出现的那只白眼雄蝇和它的红眼女儿交配,结果产生1/4红眼雄蝇、1/4红眼雌蝇、1/4白眼雌蝇、1/4白眼雄蝇,这也完全是孟德尔比数。
摩尔根根据实验结果,提出他的假设:控制白眼性状的基因W位于X染色体上,是隐性的。因为Y染色体上不带有这个基因的显性等位基因,所以最初发现的那只雄蝇()的基因型是XWY,表现为白眼,跟这只雄蝇交配的红眼雌蝇(♀)是显性基因的纯合体,基因型是++。白眼基因W是突变基因,红眼基因+是野生型基因,因为这对等位基因都在X染色体上,所以为明确起见,分别记作XW和X+,Y代表Y染色体(图1)。
白眼雄蝇与纯种红眼雌蝇交配(图1),白眼雄蝇的基因型是XWY,产生两种精子,一种精子带有X,上面有W基因,一种精子带有Y,上面没有相应的基因。红跟雌蝇的基因型是X+X+,产生的卵都带有X,上面都有一野生型基因。两种精子(XW和Y)与卵(X+)结合,子代雌蝇的基因型是X+XW,因为+对W是显性,所以表型是红眼,子代雄蝇的基因型是X+Y,所以表型也是红眼。
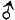
子一代的红眼雌蝇与红眼雄蝇交配时,红眼雌蝇(X+XW)产生两种卵子:一种是X+,一种是XW。红眼雄蝇也产生两种精子:一种是X+,一种是Y。卵子与精子相互结合,像图2所示那样,形成4种合子,长大后,雌蝇都是红眼(X+X+和X+XW),而雄蝇中一半是红眼(X+ Y),一半是白眼(XW Y),表型比例是2:1:1。

图2 子一代红眼雌蝇与红眼雄蝇交配,子二代雌蝇全为红眼,雄蝇中红眼和的眼各占一半
在摩尔根所做的回交试验中,子一代红眼雌蝇与白眼雄蝇交配,子一代红眼雌蝇的基因型是X+XW,产生两种卵子,一种是X+,一种是XW。白眼雄蝇的基因型是XWY,产生两种精子,一种是XW,一种是Y。雌雄配子结合后,像图3所示那样,子一代有4种表型:红眼雌蝇(X+XW)、白眼雌蝇(XW XW)、红眼雄蝇(X+Y)、白眼雄蝇(XW Y),比例是1:1:1:1。
摩尔根圆满地说明了他的实验结果,他为了验证他的假设,设计了三个新的实验:
(1)根据假设,子二代雌蝇虽然都是红眼,但基因型有两种,半数是X+ X+,半数是X+XW(图2),所以子二代雌蝇与白眼雄蝇做单对交配时,应当半数子二代雌蝇所产的后裔全部是红眼,半数子二代雌蝇则与子一代雌蝇回交一样(图3),所产的后裔是1/4红眼雌蝇:1/4白眼雌蝇:1/4红眼雄蝇:1/4白眼雄蝇。

(2)根据假设,白眼雌蝇与红眼雄蝇交配时,子代中雌蝇都是红眼,雄蝇都是白眼(图4)。
(3)根据假设,白眼雌蝇和白眼雄蝇交配时,子代雌雄都是白眼,而且以后也能真实传代,成为稳定的品系。
这三个实验中,以第二个实验最为关键,实验的结果跟预期完全符合,假设得到证实。
这样摩尔根就把决定红眼和自眼的基因定位在X染色体上。
