在最近利用RFLP标记建立的不同物种的连锁图中发现,近缘物种的RFLP标记排列次序往往是一致的,这种现象称之为共线性。但这种共线性会被倒位打断,如6个RFLP标记在番茄的一条染色体上的排列次序为 A B C D E F G ,而在同为茄科的马铃薯上的排列次序为
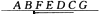 。这说明在这二个物种进化分歧时出现了C D E F的倒位。请回答以下二个问题: 1.为什么用RFLP标记可以研究近缘物种连锁图的共线性关系 2.你觉得倒位与物种进化有关吗为什么
。这说明在这二个物种进化分歧时出现了C D E F的倒位。请回答以下二个问题: 1.为什么用RFLP标记可以研究近缘物种连锁图的共线性关系 2.你觉得倒位与物种进化有关吗为什么
参考答案:
解析:1.用RFLP标记之所以可以研究近缘物种连锁图的共线性点,是因为在分子水平上,同一位点的不同等位基因有着不同的DNA序列,这种差异能以限制性内切核酸酶酶切片段长度的多态性(RFLP)表现出来。这种DNA的多态性也可能具有表型效应,但许多DNA的多态性并没有任何典型的形态学表现。用DNA印迹法,很容易鉴定出这种DNA序列差异而导致的多态性,从而得出大量的多态性标记位点。 一般情况下,RFLP与同功酶标记位点一样,是按孟德尔式共显性标记遗传的。在用RFLP标记定位时,并不需要知道有关性状的任何生化性质,也不必分离特定的基因,只要有单一顺序的标记即可。由于能够得到大量的这种DNA顺序标记,所以对不同物种cDNA来说,利用其他位点与RFLP位点的连锁分析,最终可以得出整个基因组的基因连锁图谱。 2.染色体倒位是生物染色体结构改变的重要方式,也是推动物种形成的因素之一。在自然界的昆虫群体中,特别是在果蝇属及尖眼蕈蚊属的自然群体中,曾发现大量的倒位存在,主要是臂内倒位。在鸭跖草属植物中,几乎很少个体没有倒位的。 常常有多个倒位出现在同一种群的果蝇中,形成遗传多态现象(polymorphism)。但这些倒位的分布次数随生态环境的不同而不断发生变化。例如,曾在果蝇(D. pseudoobscuria)的同一个自然群体中发现8个倒位,但它们的相对频率随地区和季节的不同而不同。在美国内华达山区的低海拔区,未倒位的标准型(ST)频率较高,一种称作矢头倒位(AR)的类型频率较低。但到高海拔区,则情况恰恰相反。另一种倒位称作CH的,出现频率不受海拔高度的影响。ST类型不管在任何海拔高度下,其频率变化都与温度相关。在温暖环境下频率较高,在冷凉环境下频率较低,而CH型则相反。这种情形可以在实验室中得到验证。从一个D. pseudoobsuna群体中取一个样本,具有11%的ST和89%的CH。在25℃下培养,5个月时检查,ST型增加了3倍。到9个月时,群体达到平衡,ST型增加到70%,而CH型下降到30%。同样的试验在16℃温度下进行,9个月时检查,ST和CH频率与原始群体未发生明显变化。 杂合倒位体与纯合体相比,具有较强的适应能力和选择上的优越性。例如,根据幼虫的生活力、发育速度、产卵量、寿命长短等指标来计算各种染色体组合在25℃温度下的适合度(fitness),假定ST/GH为1.0,那么ST/ST为0.89,而CH/CH仅为0.41。 果蝇属除D. pseudoobsuna之外,由倒位引起的遗传多态现象在其他种如D. persimilis、D. mebulosa和D. busckii中也普遍存在。在D. willistoni群体中,曾经发现了50个倒位,分布在从美国的佛罗里达至阿根廷的广大区域。在其中心区——巴西中部,平均每只果蝇具有9个倒位。 自然界物种的形成需要一定的自然隔离作用。由于染色体倒位段的存在,可以抑制交换的发生。倒位段的单交换组合常因缺失、重复而不能传递下去,这样就能使倒位段上的基因得到完整的保留,起到自然隔离的作用。一般而言,自然选择常对一段基因组团发生作用而不是对个别基因表现显著影响。倒位段内的基因组团经过后来逐渐发生而累积的随机突变,将与原来段内的基因组有很大的差异。倒位段的长度也十分重要,如果太长,有可能因双交换作用而产生基因重组,打破自然隔离。而如果太短,又没有足够的空间来积累自发突变,因而难于对自然选择起明显反应。所以自然选择往往只保留那些长短适中的倒位。 当种族间的遗传差异逐渐增加以后,不同种族间可能发生的杂交往往会产生杂种优势,这对自然选择和物种保存都是有利的。 尽管臂间倒位在自群体中比臂内倒位要少得多,但小型的臂间倒位在某些动物如黑鼠(R. attus rattus)、褐鼠(R. norvegicus)以及蚱蜢中还是存在的。它们也构成了这些生物的遗传多态现象。在真菌中的红色链孢霉,也曾经发现3个臂间倒位存在。